Bis dahin war die Gattung Alteromonas zusammen mit Pseudomonas als „Zufluchtsort“ und „Sammelbecken“ für sehr verschiedene Gram-negative, heterotrophe, aerobe Bakterien mit einer polaren (am Ende der Zellen befindlichen) Geißeln verwendet worden. Lediglich der G+C-Gehalt diente als Unterscheidungskriterium für diese beiden Gattungen: 38 bis 50 mol% für Alteromonas-Arten, 55 bis 64 mol% für Pseudomonas-Arten.
rRNA-DNA-Hybridisierungsexperimente hatten dann 1995 gezeigt, dass die so definierte Gattung Alteromonas sehr heterogen war und anhand der rRNA etwa drei Gruppen unterschieden werden konnten. Zudem unterstützte die sehr geringe Homologie der Genome der Alteromonas-Typusart A. macleodii mit allen anderen damals bekannten Arten der Gattung, dass diese Organismen nicht verwandt sind. Daher wurden die bisherigen Alteromonas-Arten bis auf diese Typusart in die neue Gattung Pseudoalteromonas übergeführt.[1]
Die Typusart der Gattung Pseudoalteromonas ist P. haloplanktis, der Referenzstamm dieser Art ist ATCC 14393 (alias Stamm 215 von Baumann et al., 1972).[2][1]
Pseudoalteromonas ist eine Gattung aerober, marinerBakterien. Die phänotypische Beschreibung der Gattung Pseudoalteromonas entspricht im Wesentlichen der Beschreibung Gattung Alteromonas: Die Zellen aller Pseudoalteromonas-Arten sind Gram-negative, nicht sporenbildende, gerade oder gekrümmte Stäbchen mit einer Länge von 1,8 bis 3,0 μm bei einem Durchmesser von 0,2 bis 1,5 μm.[1]
Die Zellen der meisten Arten sind durch einzelne, nicht umhüllte endständige (‚polare‘) Geißeln beweglich; einige Arten wie z. B. P. luteoviolacea und P. denitrificans haben jedoch umhüllte Geißeln.[1]
Es gibt auch Arten bzw. Stämme wie SM9913, die neben der polaren noch über seitliche (‚transversale‘) Geißeln verfügen.[3]
Es gibt keine Lumineszenz, aber mehrere Arten produzieren Pigmente.
Alle Arten wachsen bei ca. 20 °C, benötigen für ihr Wachstum ein Substrat auf Meerwasserbasis.
Es handelt sich um streng aerobe, chemoorganotrophe Organismen mit Atmungs-, aber nicht Gärungs-Stoffwechsel.
Es wurden Pseudoalteromonas-Vertreter identifiziert, die in der Tiefsee bei Temperaturen unter 5 °C leben.
Diese Bakterien verfügen über eine Oxidase, die Katalaseaktivität ist im Allgemeinen aber schwach und unregelmäßig.
Mit Ausnahme von P. denitrificans sind die Mitglieder der Gattung nicht zur Denitrifikation fähig.
Keiner der Stämme hat ein konstitutives Arginindihydrolase-Stoffwechselsystem.
Die Stämme akkumulieren keine Polyhydroxybuttersäure.
Viele Stämme benötigen organische Wachstumsfaktoren.
Die folgende Kombination von Eigenschaften findet sich bei allen 12 ursprünglich zugehörigen Arten von 1995:
Pseudoalteromonas-Stämme können potenziell wertvolle biologisch aktive Verbindungen, wie z. B. Proteasen,[A. 1] synthetisieren. Die Gattung ist die bedeutendste Gruppe von kultivierbaren Protease-produzierenden Meeresbakterien.[4]
Durch den Pseudoalteromonas-Stamm McH1-7 erzeugte Hemmzone (englischZone of inhibition, ZOI) der auf verschiedene andere Bakterienstämme getupft wurde. aAlteromonas sp. McT4-15, bLeisingera sp. McT4-56, cVibrio coralliilyticus OfT6-21. Balken: 10 mm.Erkrankte SteinkoralleMontastraea cavernosa, behandelt mit filtriertem Meereswasser (FSW) (a-d, 100 % Verlust nach 11 Tagen) bzw. mit Stamm McH1-7 (e-h, nur 5 % Verlust nach 11 Tagen).M. cavernosa: a-d: ein mit Stamm McH1-7 vorbehandeltes gesundes Fragment (links) in Kontakt mit einem unbehandelten gesunden Fragment (rechts), e-h: unbehandeltes gesundes Fragment (links) in Kontakt mit einem kranken Fragment (rechts) – Ansteckung und i-l ein mit McH1-7 vorbehandeltes gesundes Fragment (links) in Kontakt mit einem kranken Fragment (rechts) – signifikant weniger Verlust.
Der Pseudoalteromonas-Stamm McH1-7 gilt in der NCBI-Taxonomie als „unclassified“ mit provisorischem Artnamen Pseudoalteromonas sp. McH1-7;[7] die GTDB ordnet diesen Stamm jedoch der Spezies P. peptidolytica zu.[8]
Nach einer von Blake Ushijima, Valerie Paul et al. im Mai 2023 veröffentlichten Studie ist dieser Stamm das erste wirksame bakterielle Probiotikum, das die Korallenkrankheit Stony Coral Tissue Loss Disease (SCTLD)[9] behandeln und abwehren kann. SCTLD befällt mindestens 24 Arten von Steinkorallen, wichtige Lebensräume für Fische und andere Meerestiere mit wirtschaftlichem und ökologischem Wert, da sie dazu beitragen, die Küsten vor Sturmschäden zu schützen. Sobald eine Koralle infiziert ist, kann ihre Polypenkolonie innerhalb weniger Wochen absterben. Die schlecht verstandene Krankheit hat seit 2014 in den Korallenriffen Floridas verheerende Schäden angerichtet und breitet sich in der Karibik weiter schnell aus; Fälle von SCTLD wurden bereits in mindestens 20 Ländern bestätigt.[10]
Die neue Methode stellt nun eine vielversprechende Alternative zum bislang verwendeten BreitbandantibiotikumAmoxicillin dar. Amoxicillin ist zwar bislang (Stand 2023) das einzige Mittel zur Behandlung der Krankheit mit nachgewiesener Wirkung, birgt aber das Risiko bei den Bakterien Resistenzen zu fördern.[10]
Die schützende Wirkung von McH1-7 wurde entdeckt, als Ushijima, Paul et al. dass die Große Sternkoralle (Montastraea cavernosa) an einigen Partien schnell die charakteristischen Läsionen von SCTLD entwickelten und abstarben, während andere Stücke verschont blieben. Bei der eingehenden Untersuchung fanden sich 83 Bakterienstämme mit einer gewissen antimikrobiellen Aktivität, aber McH1-7 zeigte eine besonders deutliche Wirkung: Dieser Stamm stoppte oder verlangsamte das Fortschreiten der Krankheit bei mehr als zwei Drittel von 22 infizierten Korallenfragmenten. Mehr noch, McH1-7 verhinderte die Ausbreitung der Krankheit in allen 12 Übertragungsexperimenten und übertraf darin noch die Antibiotika.[10]
Der Stamm McH1-7 zeigt ein breites Spektrum an antibakterieller Aktivität nicht nur gegen Isolate von Bakterien, die mit SCTLD in Verbindung gebracht werden.
Nach chemische Analysen produziert McH1-7 mindestens zwei potenzielle antibakterielle Wirkstoffe, Korormicin[11] und Tetrabromopyrrol.[12]
Eine Genomanalyse identifizierte die mutmaßlichen Gene für eine L-Aminosäure-Oxidase und mehrere antibakterielle Metalloproteasen (genannt Pseudoalterine[13]).[A. 2]
McH1-7 ist damit das chemisch am besten charakterisierte Korallenprobiotikum für eine wirksame direkte und prophylaktische Behandlung für SCTLD, und damit eine mögliche Alternative zum Einsatz von Antibiotika.
Neben diesen marinen Zielen scheint auch eine Wirkung gegen terrestrische Ziele (d. h. auf dem Land), darunter auch menschliche Krankheitserreger gegeben oder möglich zu sein; sowohl Gram-positive als auch Gram-negative.[10]
Zwar sind nützliche korallenassoziierte Bakterien im Vergleich zu den für das menschliche Darmmikrobiom geeigneten Probiotika noch wenig erforscht. Diese Studien deuten jedoch darauf hin, dass probiotische Bakterien eine praktikable Option zur Eindämmung von Korallenerkrankungen sind.[10]
Der Pseudoalteromonas-Stamm PAMC 21717 wird der Art P. arctica zugeordnet.
Er produziert die Protease Pro21717, das als Waschmittelenzym Anwendung findet.[10]
Aus Organismen, die in kalten Lebensräumen vorkommen, isolierte Enzyme weisen im Allgemeinen eine höhere katalytische Aktivität bei niedrigen Temperaturen auf als ihre Homologe aus mesophilen Organismen in gemäßigten Zonen.
Unter diesen kälteaktiven Enzymen sind die Proteasen für eine Reihe biotechnologischer Anwendungen sehr nützlich. Dazu gehören insbesondere Wirkstoffe in Wasch- und Geschirrspülmitteln, da sie bereits in kaltem oder mäßig warmem Wasser eine starke proteinabbauende Aktivität aufweisen.[15]
Ha Ju Park et al. identifizierten in dem kälteverträglichen (psychrophilen) Pseudoalteromonas-Stamm PAMC 21717 eine solche als Pro21717 bezeichnete kälteaktive Protease und bestimmten die Struktur ihrer katalytischen Domäne (CD) mit einer Auflösung von 0,14 nm. Die CD-Struktur von Pro21717 zeigt eine Faltung ähnlich wie bei der Protease Subtilisin. Verschiedene Parameter lieferten zudem mögliche Erklärungen für die kälteangepassten Eigenschaften von Pro21717.
Diese Ergebnisse könnten eine Grundlage für industrielle Anwendungen von Pro21717 als kälteaktives Wasch- oder Geschirrspülmittelenzym bilden.[15]
Produktion von Geißeln und Schwarmmotilität des Pseudoalteromonas-Stammes SM9913, Wildtyp und Mutante ΔfilZ. Details siehe Sheng et al. (2023)[3]Einfluss des Proteins FilZ auf die Schwarmmotilität des Pseudoalteromonas-Stammes SM9913, beim Wildtyp und verschiedenen Mutanten. Details siehe Sheng et al. (2023)[3]
Der Pseudoalteromonas-Stamm SM9913 gilt in der NCBI-Taxonomie als „unclassified“ mit provisorischem Artnamen Pseudoalteromonas sp. SM9913;[7] die GTDB ordnet diesen Stamm jedoch der Spezies P. tetraodonis zu.[8]
Bei dem Stamm SM9913 handelt es sich um ein Bakterium der Tiefsee-Sedimente.
Der Stamm besitzt (wie auch einige andere Bakterien) die gattungstypische immer vorhandene (‚konstitutive‘) endständige (‚polare‘) Geißel, kann aber auch ggf. durch bestimmte Umgebungsbedingungen induzierbare seitliche (‚laterale‘) Geißeln ausbilden.
Diese Bakterien können daher sowohl im freien Wasser schwimmen, als auch auf viskosen Unterlagen wie dem Tiefseesediment schwärmen.[A. 3]
Die konstitutive polare Geißel ist für das Schwimmen, die induzierten lateralen Geißeln für das Schwärmen unerlässlich.
Dies ist eine spezielle Anpassung an die extremen Bedingungen der Tiefseesedimente als spezieller mikrobieller Lebensraum.[3]
Die Steuerung des Schwarmverhaltens ist bei begeißelten Bakterien kompliziert, insbesondere wenn sie wie SM9913 ein duales Geißelsystem besitzen.
Die beiden Geißelsysteme werden, wiei von Qi Sheng et al. 2023 beschrieben, von zwei unabhängigen Genclusternkodiert. [23,26]. Die Funktion des polaren Flagellums wird durch Chemotaxis vermittelt. Der Stamm SM9913 hat aufgrund einer Nonsense-Mutation im Flagellin-Gen der polaren Gencluster eine im Vergleich zu anderen Pseudoalteromonas-Stämmen abnorm kurze polare Geißel (Flagellum). Ds kurze polare Flagellum hat offenbar nur eine geringe Auswirkung auf die Schwimmfähigkeit des Stammes,
auch wenn die Autoren nicht ausschließen können, dass sich die Bakterien dieses Stamms im bestimmten Situationen sich etwas anders verhalten könnten als andere Vertreter der Gattung mit normal langem polaren Flagellum.[3]
Die Bewegung der polaren Geißel wird durch ein FilZ genanntes Effektor-Protein herunterreguliert, das selbst im Gen filZ innerhalb des Genclusters der lateralen Flagellen kodiert ist. Die Funktion von FilZ ihrerseits wird durch intrazelluläres Cyclo-di-Guanosinmonophosphat (c-di-GMP)[16] gehemmt. Bioinformatische Untersuchungen ergaben, dass filZ-ähnliche Gene in vielen Bakterien mit dualen Geißelsystemen vorhanden sind.[3]
Der Gattungsname leitet sich ab von altgriechischψευδήςpseudḗs, deutsch ‚falsch‘ und dem Gattungsnamen Alteromonas, der sich seinerseits zusammensetzt aus lateinischalter‚ein anderer‘, ‚anders‘ und monas‚Monade‘, ‚Einheit‘ (Einzeller). Die Bezeichnung deutet also an, dass es sich um nicht echte Alteromonas-Einzeller handelt.[17]
Das Art-Epitheton der Typusspezies, haloplanktis, stammt von altgriechisch ἅλςhals, deutsch ‚Salz‘, ‚Meer‘ (Genitiv: halos), und πλαγκτόςplanktos, deutsch ‚das Umherirrende‘, ‚Umherstreifende‘ oder ‚Wandernde‘.[17] Die Bezeichnung deutet an, dass es sich um im Meer bzw. Salzwasser umhergetriebene oder -wandernde Organismen handelt (vgl. Plankton).
Die Gattung wurde 1995 von G. Gauthier et al. 1995 als Abspaltung von Alteromonas neu eingerichtet mit den ursprünglichen Spezies (Arten):
P. atlantica, P. aurantia, P. carrageenovora, P. citrea, P. denitrificans, P. espejiana, P. haloplanktis (Typusart, damals mit zwei Subspezies, P. h. subsp. haloplanktis und P. h. subsp. tetraodonis), P. luteoviolacea, P. nigrifaciens, P. rubra, P. undina und P. piscicida.[1]
Die Zuordnung der Stämme auf die einzeln Arten der Gattung ist schwierig und in etlichen Fällen noch in der Diskussion.
Die Artenliste bzw. innere Systematik der Gattung ist (mit Stand 5. September 2023) wie folgt:
In der GTDB werden die einige Stämme etwas anders auf die Arten der Gattung aufgeteilt, wodurch einerseits unter den Artnamen Synonymien entstehen, andererseits aber (durch angehängte Großbuchstaben gekennzeichnete) Abspaltungen auftreten; insbesondere sind die Stämme der LPSN-Art P. fuliginea in der GTDB komplett auf andere Arten verteilt:
PseudoalteromonasGauthieret al. 1995 (L) bzw.
Gauthieret al. 1995 emend. Beurmannet al. 2017[18] (N) mit Schreibvariante Pseudalteromonas (L)
Pseudoalteromonas aestuariivivensParket al. 2016 (L,N) inkl. P. sp. DB-2 (N)
Pseudoalteromonas agarivoransRomanenkoet al. 2003 (L,N: G: Syn. von P. atlantica)
Pseudoalteromonas nigrifaciens (Beurmannet al. 1984 ex White 1940) Gauthieret al. 1995 (G), veraltet Alteromonas nigrifaciens (ex White 1940) Beurmannet al. 1984 mit Schreibvariante Alteromonas nigrificans[27]
Referenzstamm: 217 alias ATCC:19375, CIP:104111, DSM:8810, IAM:13010, JCM:20792, KMM:661, BCCM/LMG:2227, NBRC:103036, NCIMB:8614 oder NCTC:10691 (N)
Pseudoalteromonas nigrifaciens (Beurmannet al. 1984 ex White 1940) Gauthieret al. 1995 (L,N) ⇒ Alteromonas nigrifaciens (ex White 1940) Beurmannet al. 1984 (nicht bei G)
Zu einigen der Arten sind Referenzstamm (Typstamm, oft gekennzeichnet durch ein hochgestelltes T) und andere Stämme angegeben. Wie stets ist der Doppelpunkt im Namen der Stämme einem Leerzeichen äquivalent.
Siphovirale und filamentöse PSA-Phagen sind in den Polargebieten vorherrschend, während einige myovirale und siphovirale PSA-Phagen in der Tiefsee häufiger vorkommen.[38]
Die Viren oder Phagen dieser Klasse haben meist ein sehr enges Wirtsspektrum und infizieren gewöhnlich nur Bakterien (oder ggf. Archaeen) einer bestimmten Art oder gar innerhalb dieser nur eines bestimmen Stammes; das gilt auch für die PSA-Phagen dieser Gruppe. Unter den PSA-Phagen sind alle drei in dieser Klasse bekannten Morphotypen vertreten: Siphoviren, Myoviren und Podoviren (diese wurden früher als – sehr große – Virusfamilien angesehen).
Genom-Vergleich der Viruspartikel verschiedener Viren aus der Klasse Caudoviricetes (Phagen mit Kopf-Schwanz-Aufbau): Shewanella-Phage 1/44 (Morphotyp der Siphoviren), Enterobacteria-Phage Mu (Muvirus mu, Morphotyp der Myoviren) und die chimären Beispiele PSA-HS6 und Pseudomonas-Phage D3112 (Casadabanvirus D3112) mit Myo-Kopf und Sipho-Schwanz.
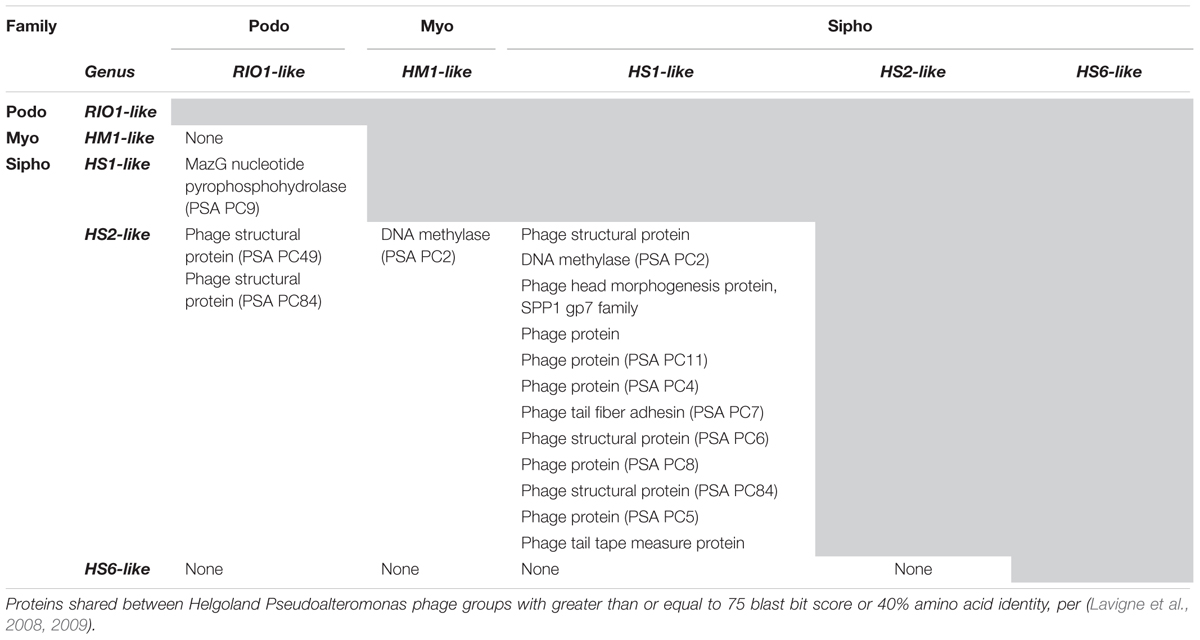
Interessanterweise zeigen einige der sog. Helgoland-Pseudoalteromonas-Phagen[A. 7]hybride Merkmale aus diesen drei Caudoviricetes-Morphotypen in sich vereinigt, darunter die Phagen HS1, HS2 und HS6.
Beispielsweise sind die HS6-ähnlichen Phagen modulare Hybride, die aus einem Siphovirus-ähnlichen, nicht kontraktilen „Schwanz“ bestehen, der mit einem Myovirus-Mu-ähnlichen „Kopf“ (Kapsid) verschmolzen ist.[43] Die folgenden Zuordnungen zu Morphotypen sind daher eher tentativ zu verstehen:
Zi-Chao Yu et al. isolierten 2015 den fadenförmigen (filamentösen) Phagen f327 aus dem Pseudoalteromonas-Stamm BSi20327 vom arktischen Meereis. Sie zeigten auch, dass diese Gruppe von Phagen im arktischen Meereis weit verbreitet ist. Eine Bestätigung durch das ICTV steht derzeit (September 2023) noch aus:[70]
Pseudoalteromonas filamentous phage f327, Wirt: P. sp. BSi20327
Es gibt derzeit (Stand August 2023) Metagenomik-Hinweise auf 16 weitere filamentöse PSA-Phagen (darunter PCG2, PCG6, PCG11, PCG15 und PCG20) und nochmals so viele ins Wirtsgenom integrierte Prophagen.[38]
↑Proteasen sind Enzyme, die Peptidbindungen von Proteinen aufbrechen (hydrolysieren).
↑Pseudoalterine werden auch von anderen Pseudoalteromonas-Stämmen wie CF6-2 produziert.[14]
↑Bei den Fortbewegungsarten von Bakterien unterscheidet man zwischen Schwimmen (englischswimming) und Schwärmen (engl. swarming ). Schwimmen ist die von einer oder mehreren rotierenden Geißeln angetriebene Bewegung einzelner Zellen in wässriger Umgebung; Schwärmen ist die ebenfalls von Geißeln angetriebene schnelle Bewegung einer großen Anzahl von Zellen auf viskosen Oberflächen.
↑Die folgende Einteilung in Unterarten (Subspezies) ist mit der Hochstufung von P. haloplanktis subsp. tetradontis zu einer eigenen Spezies obsolet geworden:
↑die Zuordnung zum Morphotyp Podoviren ist noch nicht sicher (Referenz Rahlff 2023 ist bisher (Stand 9. September 2023) nur Preprint)
↑Zum NCBI-gelisteten Pseudoalteromonas-Phage PSA-HS9 konnte keine Primärquelle gefunden werden, umgekehrt ist der bei Duhaime (2017) beschriebene Phage HS8 nicht in der NCBI-Taxonomie. Möglicherweise liegt ein Verschreiber vor und es sollte immer HS8 heißen.
↑Pseudoalteromonas-Phage KB12-38 ist eng verwandt mit Achromobacter-Phage JWF
↑ abcdefghij
G. Gauthier, Michel J. Gauthier, Richard Christen: Phylogenetic Analysis of the Genera Alteromonas, Shewanella, and Moritella Using Genes Coding for Small-Subunit rRNA Sequences and Division of the Genus Alteromonas into Two Genera, Alteromonas (Emended) and Pseudoalteromonas gen. nov., and Proposal of Twelve New Species Combinations. In: Microbiology Society: International Journal of Systematic Bacteriology, Band 45, Nr. 4, Oktober 1995, S. 755–761, ISSN1466-5034; OCLC43894812, doi:10.1099/00207713-45-4-755, PMID 7547295 (englisch).
↑ abcdefghi
Qi Sheng, Ang Liu, Peiling Yang, Zhuowei Chen, Peng Wang, Haining Sun, Chunyang Li, Andrew McMinn, Yin Chen, Yuzhong Zhang, Hainan Su, Xiulan Chen, Yuqiang Zhang: The FilZ Protein Contains a Single PilZ Domain and Facilitates the Swarming Motility of Pseudoalteromonas sp. SM9913. In: MDPI: Microorganisms, Band 11, Nr. 6, Section Environmental Microbiology, 13. Juni 2023, S. 1566; doi:10.3390/microorganisms11061566 (englisch).
↑ abcd
Dan Liu, Huang Jiafeng, Cuiling Wu, Congling Liu, Ran Huang, Weng Wang, Tingting Yin, Xiaotao Yan, Hailun He, Leilei Chen: Purification, Characterization, and Application for Preparation of Antioxidant Peptides of Extracellular Protease from Pseudoalteromonas sp. H2. In: MDPI: Molecules, Band 24, Nr. 16, Section Chemical Biology, S. 3373; doi:10.3390/molecules24183373, ResearchGate (englisch).
↑ abcdefghijk
Blake Ushijima, Sarath P. Gunasekera, Julie L. Meyer, Jessica Tittl, Kelly A. Pitts, Sharon Thompson, Jennifer M. Sneed, Yousong Ding, Manyun Chen, L. Jay Houk, Greta S. Aeby, Claudia C. Häse, Valerie J. Paul: Chemical and genomic characterization of a potential probiotic treatment for stony coral tissue loss disease. In: Nature: Communications Biology, band 6, Nr. 248, 6. April 2023; doi:10.1038/s42003-023-04590-y. Dazu:
↑ ab
Jie Yang, Yang Yu, Bai-Lu Tang, Shuai Zhong, Mei Shi, Bin-Bin Xie, Xi-Ying Zhang, Bai-Cheng Zhou, Yu-Zhong Zhang, Xiu-Lan Chen: Pilot-Scale Production and Thermostability Improvement of the M23 Protease Pseudoalterin from the Deep Sea Bacterium Pseudoalteromonas sp. CF6-2. In: MDPI: Molecules, Band 21, Nr. 11, 17. November 2016, S. 1567; doi:10.3390/molecules21111567 (englisch).
↑ abc
Ha Ju Park, Chang Woo Lee, Dockyu Kim, Hackwon Do, Se Jong Han, Jung Eun Kim, Bon-Hun Koo, Jun Hyuck Lee, Joung Han Yim: Crystal structure of a cold-active protease (Pro21717) from the psychrophilic bacterium, Pseudoalteromonas arctica PAMC 21717, at 1.4 Å resolution: Structural adaptations to cold and functional analysis of a laundry detergent enzyme. In: PLOS ONE, Band 13, Nr. 2, 12. Februar 2018, S. e0191740; doi:10.1371/journal.pone.0191740, PMID 29466378, PMC 5821440 (freier Volltext) (englisch).
↑ abcdJ.P. Euzéby: Pseudoalteromonas. In: List of Prokaryotic names with Standing in Nomenclature (LPSN). 13. November 2015, abgerufen am 17. Dezember 2015.
↑
Silvia Beurmann, Blake Ushijima, Christina M. Svoboda, Patrick Videau, Ashley M. Smith, Stuart P. Donachie, Greta S. Aeby, Sean M. Callahan: Pseudoalteromonas piratica sp. nov., a budding, prosthecate bacterium from diseased Montipora capitata, and emended description of the genus Pseudoalteromonas." In: International Journal of Systematic and Evolutionary Microbiology, Band 67, Nr. 8, 1. August 2017, S. 2683–2688; doi:10.1099/ijsem.0.001995, PMID 28792373 (englisch).
↑ ab
Xue Meng, Min Wang, Siyuan You, Duobing Wang, Yan Li, Zhaoyang Liu, Yu Gao, Lu Liu, Yaoyuan Zhang, Zhenghao Yan, Chunyan Liu, Yong Jiang, Hongbing Shao: Characterization and Complete Genome Sequence of a Novel Siphoviridae Bacteriophage BS5. In: Current Microbiology, Band 74, 20. April 2017, S. 815–820; doi:10.1007/s00284-017-1221-2 (englisch).
↑ abc
Qi Wang, Yundan Liu, Qian Liu, Xinxin Liu, Fan Yang, Min Wang, Hongbing Shao, Yong Jiang: Isolation and Complete Genome of the Marine Pseudoalteromonas Phage C7 from Coastal Seawater of Yellow Sea, China. In: Current Microbiology, Band 77, 23. November 2019, S. 279–285; doi:10.1007/s00284-019-01815-4.
↑ abcd
Hanna M. Kivelä, Rimantas Daugelavičius, Riina H. Hankkio, Jaana K. H. Bamford, Dennis H. Bamford: Penetration of Membrane-Containing Double-Stranded-DNA Bacteriophage PM2 into Pseudoalteromonas Hosts. In: Journal of Bacteriology, Band 186. Nr. 16, August 2004, S. 5342–5354; doi:10.1128/JB.186.16.5342-5354.2004, PMID 15292135, PMC 490941 (freier Volltext) (englisch).
↑ ab
Usio Simidu, Kumiko Kita-Tsukamoto, Takeshi Yasumoto, Mari Yotsu: Taxonomy of Four Marine Bacterial Strains That Produce Tetrodotoxin. In: International Journal of Systematic and Evolutionary Microbiology, Band 40, Nr. 4, 1. Oktober 1990; doi:10.1099/00207713-40-4-331, PMID 2275851 (englisch).
↑
Kasthuri Venkateswaran, N. Dohmoto: Pseudoalteromonas peptidolytica sp. nov., a novel marine mussel-thread-degrading bacterium isolated from the Sea of Japan. In: International Journal of Systematic and Evolutionare Microbiology, Band 50, Nr. 2, 1. März 2000, S. 565–574; doi:10.1099/00207713-50-2-565, PMID 10758862, ResearchGate (englisch). Siehe insbes. Fig. 1.
↑ ab
Hakdong Shin, Ju-Hoon Lee, Chi Sang Ahn, Sangryeol Ryu, Byung Cheol Cho: Complete genome sequence of marine bacterium Pseudoalteromonas phenolica bacteriophage TW1. In: Archives of Virology, Band 159, Nr. 1, Januar 2014, S. 159-162; doi:10.1007/s00705-013-1776-6, PMID 23851651, Epub 13. Juli 2013 (englisch).
↑ ab
Melissa Beth Duhaime, Antje Wichels, Jost Waldmann, Hanno Teeling, Frank Oliver Glöckner: Ecogenomics and genome landscapes of marine Pseudoalteromonas phage H105/1. In: Nature: The ISME Journal, Band 5, Evolutionary genetics, 8. Juli 2010, S. 107–121; doi:10.1038/ismej.2010.94 (englisch). Mit Fotos der Viruspartikel.
↑ ab
René Kallies, Bärbel Kiesel, Matthias Schmidt, Nawras Ghanem, Jakob Zopfi, Jörg Hackermüller, Hauke Harms, Lukas Y. Wick, Antonis Chatzinotas: Complete genome sequence of Pseudoalteromonas virus vB_PspP-H6/1 that infects Pseudoalteromonas sp. strain H6. In: Marine Genomics,
Band 47, Oktober 2019, S. 100667; doi:10.1016/j.margen.2019.03.002 (englisch).
↑ ab
Elena Lara, Karin Holmfeldt, Natalie Solonenko, Elisabet Laia Sà, J. Cesar Ignacio-Espinoza, Francisco M. Cornejo-Castillo, Nathan C. Verberkmoes, Dolors Vaqué, Matthew B. Sullivan, Silvia G. Acinas: Life-style and genome structure of marine Pseudoalteromonas siphovirus B8b isolated from the northwestern Mediterranean Sea. In: PLOS ONE, Band 10, Nr. 1, 14. Januar 2015, S. e0114829; doi:10.1371/journal.pone.0114829, PMID 25587991, PMC 4294664 (freier Volltext) (englisch).
↑ abcdefg
Kaiyang Zheng, Yue Dong, Yantao Liang, Yundan Liu, Xinran Zhang, Wenjing Zhang, Ziyue Wang, Hongbing Shao, Yeong Yik Sung, Wen Jye Mok, Li Lian Wong, Andrew McMinn, Min Wang: Genomic diversity and ecological distribution of marine Pseudoalteromonas phages. In: Marine Life Science & Technology, Band 5, 20. Januar 2023, S. 271–285; doi:10.1007/s42995-022-00160-z (englisch). Dazu Korrektur doi:10.1007/s42995-023-00166-1.
↑
Natalya Yutin, Mike Rayko, Dmitry Antipov, Pascal Mutz, Yuri I. Wolf, Mart Krupovic, Eugene V. Koonin: Varidnaviruses in the Human Gut: A Major Expansion of the Order Vinavirales. In: MDPI: Viruses, Band 14, Nr. 9, Section Bacterial Viruses, 23. August 2022, S. 1842; doi:10.3390/v14091842 (englisch). Siehe insbes. Fig. 3. Preprint auf bioRxiv: doi:10.1101/2022.07.18.500477 (Fig. 3 in Semigraphik).
↑ abcdefghijklmn
Melissa B. Duhaime, Natalie Solonenko, Simon Roux, Nathan C. Verberkmoes, Antje Wichels, Matthew B. Sullivan: Comparative Omics and Trait Analyses of Marine Pseudoalteromonas Phages Advance the Phage OTU Concept. In: Frontiers in Microbiolog, Band 8, Sec. Virology, 6. Juli 2017, S. 1241; doi:10.3389/fmicb.2017.01241, PMID 28729861, PMC 5498523 (freier Volltext) (englisch). Siehe insbes. Tbl. 1 und Tbl. 2.
↑ abcde
Cristina Howard-Varona, Morgan Marie Lindback, Eric Bastien, Natalie Solonenko, Ahmed Zayed, Ho Bin Jang, Bill Andreopoulos, Heather M. Brewer, Tijana Glavina del Rio, Joshua N. Adkins, Subhadeep Paul, Matthew B. Sullivan, Melissa B. Duhaime: Phage-specific metabolic reprogramming of virocells. In: Nature: The ISME Journal, Band 14, Nr. 4, Januar 2020, S. 1-15; doi:10.1038/s41396-019-0580-z, ResearchGate (englisch). Siehe insbes. Fig. 1.
↑
Janina Rahlff, Matthias Wietz, Helge-Ansgar Giebel, Oliver Bayfield, Emelie Nilsson, Kristofer Bergström, Kristopher Kieft, Karthik Anantharaman, Mariana Ribas-Ribas, Oliver Wurl, Matthias Hoetzinger, Alfred Antson, Karin Holmfeldt: Ecogenomics reveals distinctive viral-bacterial communities in the surface microlayer of a natural surface slick. Auf: bioRxiv vom Februar 2023 (Preprint); doi:10.1101/2023.02.24.528798, ResearchGate (englisch).
↑
Zhaoyang Liu, Huifang Li, Min Yang, Yong Jiang, Qingwei Yang, Xinhao Zhou, Zheng Gong, Qian Liu, Hongbing Shao: Isolation, characterization and genome sequencing of the novel phage SL25 from the Yellow Sea, China. In: Marine Genomics, Band 37, Februar 2018, S. 31-34; doi:10.1016/j.margen.2017.09.008 (englisch).
↑
René Kallies, Bärbel Kiesel, Matthias Schmidt, Johannes Kacza, Nawras Ghanem, Anja Narr, Jakob Zopfi, Lukas Y. Wick, Jörg Hackermüller, Hauke Harms, Antonis Chatzinotas: Complete genome sequence of Pseudoalteromonas phage vB_PspS-H40/1 (formerly H40/1) that infects Pseudoalteromonas sp. strain H40 and is used as biological tracer in hydrological transport studies. In: BMC: Environmental Microbiome (Stand Genomic Sci), Band 12, Nr. 20, 2. Februar 2017; doi:10.1186/s40793-017-0235-5, PMID 28168014, PMC 5288847 (freier Volltext) (englisch).
↑ ab
Jianhua Sun, Xinran Zhang, Yantao Liang, Kaiyang Zheng, Fraser Kennedy, Meiaoxue Han, Gang Liu, Yundan Liu, Ziyue Wang, Xuechao Chen, Yeong Yik Sung, Wen Jye Mok, Li Lian Wong, Andrew McMinn, Min Wang: Abundance and ecological footprint of Pseudoalteromonas phage vB_PhoS_XC in the Ulva prolifera green tide. In: Frontiers in Marine Science, Band 10, 18. Juli 2023, Sec. Microbial Symbioses, S. 1201434; doi:10.3389/fmars.2023.1201434 (englisch).




























{kind=link}